45. Кора больших полушарий, ее строение и значение.
Кора больших полушарий головного мозга - структура головного мозга, слой серого вещества толщиной 1,3-4,5 мм, расположенный по периферии полушарий большого мозга, и покрывающий их.
Кора головного мозга играет очень важную роль в осуществлении высшей нервной (психической) деятельности.
У человека кора составляет в среднем 44% от объёма всего полушария в целом.
Кора большого мозга покрывает поверхность полушарий и образует большое количество различных по глубине и протяжённости борозд. Между бороздами расположены различной величины извилины большого мозга.
В каждом полушарии различают следующие поверхности:
выпуклую верхнелатеральную поверхность , примыкающую к внутренней поверхности костей свода черепа
нижнюю поверхность , передние и средние отделы которой располагаются на внутренней поверхности основания черепа, в области передней и средней черепных ямок, а задние - на намёте мозжечка
медиальную поверхность , направленную к продольной щели мозга.
В каждом полушарии различают наиболее выступающие места: спереди - лобный полюс, сзади - затылочный, и сбоку - височный.
Полушарие разделено на пять долей. Четыре из них примыкают к соответствующим костям свода черепа:
Лобная, теменная, затылочная, височная, островковая доля отделяет лобную долю от височной.
Устройство коры головного мозга и взаимодействие между собой отдельных её частей называется архитектоникой коры большого мозга. Место, где кора мозга выполняет те или иные функции: анализ поступающей от органов чувств информации, их сохранение, и т.д., во многом определяются внутренней структурой и построением связей (морфологией) внутри конкретных участков головного мозга (такие участки называются корковыми полями). Ещё одной важнейшей функцией коры головного мозга является связь с определенными внешними приемниками информации (рецепторами), каковыми являются все органы чувств, а также с органами и тканями, выполняющими команды, поступающие от коры головного мозга (эффекторами).
Всё, что человек видит, распознаётся и анализируется в затылочной области коры головного мозга, глаз же - всего лишь приемник изображения, передающий его по нервным волокнам для анализа в затылочную зрительную зону.
В том случае, если изображение движется, то анализ движения этого изображения происходит в теменной области , и в результате этого анализа мы определяем, в каком направлении и с какой скоростью движется видимый нами предмет.
Теменные области коры, совместно с височными зонами коры принимают участие в формировании акта членораздельной речи и в восприятии формы тела человека и его расположения в пространстве.
Лобные доли коры головного мозга у человека являются теми участками коры, которые в основном осуществляют высшие психические функции, проявляющиеся в формировании личностных качеств, темперамента, характера, способностей, воли, разумности поведения, творческих наклонностей и одарённости, влечений и пристрастий, в общем всего того, что делает человека личностью, не похожей на всех остальных людей, и в построении целенаправленного поведения, основанного на предвидении. Все эти способности резко нарушается при повреждении лобных отделов коры головного мозга.
Наиболее обширное поражение коры головного мозга сопровождается полным исчезновением психической деятельности.
глиальные клетки ; оно расположено в некоторых отделах глубинных мозговых структур, из этого вещества сформирована кора больших полушарий (а также мозжечка).Каждое полушарие разделяется на пять долей, четыре из которых (лобная, теменная, затылочная и височная) примыкают к соответствующим костям черепного свода, а одна (островковая) находится в глубине, в ямке, которая разделяет лобную и височную доли.
Кора большого мозга имеет толщину в 1,5–4,5 мм, ее площадь увеличивается за счет присутствия борозд; она связана с другими отделами ЦНС, благодаря импульсам, которые проводят нейроны.
Полушария достигают примерно 80% от общей массы головного мозга. Они осуществляют регуляцию высших психических функций, тогда как мозговой ствол – низшие, которые связаны с деятельностью внутренних органов.
Три основные области выделяют на полушарной поверхности :
- выпуклая верхнелатеральная, которая примыкает к внутренней поверхности черепного свода;
- нижняя, с располагающимися передними и средними отделами на внутренней поверхности черепного основания и задними в области намета мозжечка;
- медиальная расположена у продольной щели мозга.
Особенности устройства и деятельности
Кора большого мозга подразделяется на 4 вида:
- древняя – занимает чуть более 0,5% всей поверхности полушарий;
- старая – 2,2%;
- новая – более 95%;
- средняя – примерно 1,5%.

Филогенетически древняя кора большого мозга, представленная группами крупных нейронов, оттесняется новой к основанию полушарий, становясь узкой полоской. А старая, состоящая из трех клеточных слоев, смещается ближе к середине. Главная область старой коры – гиппокамп, являющийся центральным отделом лимбической системы . Средняя (промежуточная) кора представляет собой образование переходного типа, так как трансформация старых структур в новые осуществляется постепенно.
Кора головного мозга у человека, в отличие от таковой у млекопитающих, также ответственна за согласованную работу внутренних органов. Такое явление, при котором, возрастает роль коры в осуществлении всей функциональной деятельности организма, носит название кортикализация функций.
Одна из особенностей коры – ее электрическая активность, происходящая спонтанно. Нервные клетки, расположенные в этом отделе, обладают определенной ритмической активностью, отражающей биохимические, биофизические процессы. Активность обладает различной амплитудой и частотой (альфа-, бета-, дельта-, тета-ритмы), что зависит от влияния многочисленных факторов (медитации, фазы сна, переживания стресса, наличия судорог, новообразования).
Структура
Кора головного мозга представляет собой многослойное образование: каждый из слоев имеет свой определенный состав нейроцитов, конкретную ориентацию, расположение отростков.
Систематическое положение нейронов в коре носит название «цитоархитектоника», расположенные в определенном порядке волокна – «миелоархитектоника».
Кора больших полушарий головного мозга состоит из цитоархитектонических шесть слоев.

- Поверхностный молекулярный, в котором нервных клеток не очень много. Их отростки расположены в нем самом, и они не выходят за пределы.
- Наружный зернистый сформирован из пирамидальных и звездчатых нейроцитов. Отростки выходят из этого слоя и идут в последующие.
- Пирамидальный состоит из пирамидных клеток. Их аксоны направляются вниз, где оканчиваются или формируют ассоциативные волокна, а дендриты идут вверх, во второй слой.
- Внутренний зернистый образован звездчатыми клетками и малыми пирамидными. Дендриты идут в первый слой, боковые отростки разветвляются в пределах своего слоя. Аксоны протягиваются в верхние слои или в белое вещество.
- Ганглионарный образован большими пирамидными клетками. Здесь находятся самые крупные нейроциты коры. Дендриты направлены в первый слой или распределены в своем. Аксоны выходят из коры и начинают являться волокнами, связывающими различные отделы и структуры ЦНС между собой.
- Мультиформный – состоит из различных клеток. Дендриты идут к молекулярному слою (некоторые только до четвертого или пятого слоев). Аксоны направляются в вышележащие слои или выходят из коры в качестве ассоциативных волокон.
Кора головного мозга разделяется на области – так называемая горизонтальная организация . Всего их насчитывается 11, и они включают в себя 52 поля, каждое из которых имеет свой порядковый номер.
Вертикальная организация
Существует и вертикальное разделение – на колонки нейронов. При этом маленькие колонки объединяются в макроколонки, которые называют функциональным модулем. В основе таких систем находятся звездчатые клетки – их аксоны, а также горизонтальные связи их с боковыми аксонами пирамидальных нейроцитов. Все нервные клетки вертикальных колонок реагируют на афферентный импульс одинаково и вместе посылают эфферентный сигнал. Возбуждение в горизонтальном направлении обусловлено деятельностью поперечных волокон, которые следуют от одной колонки к другой.
Впервые обнаружил единицы, которые объединяют нейроны различных слоев по вертикали, в 1943г. Лоренте де Но – с помощью гистологии. Впоследствии это было подтверждено с помощью методов электрофизиологии на животных В. Маунткаслом.

Развитие коры во внутриутробном развитии начинается рано: уже в 8 недель у эмбриона появляется корковая пластина. Вначале дифференцируются нижние слои, а в 6 месяцев у будущего ребенка появляются все поля, которые присутствуют и у взрослого человека. Цитоархитектонические особенности коры к 7 годам полностью формируются, но тела нейроцитов увеличиваются еще до 18. Для образования коры необходимо согласованное перемещение и деление клеток-предшественниц, из которых появляются нейроны. Установлено, что на этот процесс влияет специальный ген.
Горизонтальная организация
Принято разделять зоны коры головного мозга на:
- ассоциативные;
- сенсорные (чувствительные);
- моторные.
Учеными при изучении локализованных участков и их функциональных особенностей применялись разнообразные способы: раздражение химическое или физическое, частичное удаление мозговых участков, выработка условных рефлексов, регистрация биотоков мозга.
Чувствительные
Эти области занимают примерно 20% коры. Поражение таких зон ведет к нарушению чувствительности (снижение зрения, слуха, обоняния и т. п.). Площадь зоны напрямую зависит от количества нервных клеток, которые воспринимают импульс от определенных рецепторов: чем их больше, тем выше сензитивность. Выделяют зоны:
- соматосенсорную (отвечает за кожную, проприоцептивную, вегетативную чувствительность) – она расположена в теменной доле (постцентральная извилина);
- зрительную, двухстороннее повреждение которое приводит к полной слепоте, – находится в затылочной доле;
- слуховую (расположена в височной доле);
- вкусовую, находящуюся в теменной доле (локализация – постцентральная извилина);
- обонятельную, двухстороннее нарушение которой приводит к потере обоняния (расположена в гиппокамповой извилине).
Нарушение слуховой зоны не приводит к глухоте, но появляются другие симптомы. Например, невозможность различения коротких звуков, смысла бытовых шумов (шагов, льющейся воды и т. п.) при сохранности различия звуков по высоте, длительности, тембру. Также может происходить амузия, заключающаяся в неспособности узнавать, воспроизводить мелодии, а также различать их между собой. Музыка также может сопровождаться неприятными ощущениями.
Импульсы, идущие по афферентным волокнам с левой стороны тела, воспринимаются правым полушарием, а с правой стороны – левым (повреждение левого полушария вызовет нарушение чувствительности с правой стороны и наоборот). Это связано с тем, что каждая постцентральная извилина связана с противоположной частью тела.
Двигательные
Моторные участки, раздражение которых вызывает движение мускулатуры, располагаются в передней центральной извилине лобной доли. Двигательные зоны сообщаются с сенсорными.
Двигательные пути в продолговатом мозге (и частично в спинном) образуют перекрест с переходом на противоположную сторону . Это приводит к тому, что раздражение, которое возникает в левом полушарии, поступает в правую половину туловища, и наоборот. Поэтому поражение участка коры одного из полушарий ведет к нарушению двигательной функции мышц с противоположной стороны туловища.
Моторная и сенсорная области, которые расположены в районе центральной борозды, объединяются в одно образование – сенсомоторную зону.
Неврология и нейропсихология накопили множество сведений о том, как поражение этих областей приводит не только к элементарным двигательным расстройствам (параличам, парезам, треморам), но и к нарушениям произвольных движений и действий с предметами – апраксиям. При их появлении могут нарушаться движения во время письма, происходить расстройства пространственных представлений, появляться бесконтрольные шаблонные движения.
Ассоциативные
Эти зоны ответственны за связывание поступающей сенсорной информации с той, которая была получена ранее и хранится в памяти. Кроме того, они позволяют сравнивать между собой информацию, которая идет от различных рецепторов. Ответная реакция на сигнал формируется в ассоциативной зоне и передается в зону двигательную. Таким образом, каждая ассоциативная область отвечает за процессы памяти, научения и мышления . Крупные ассоциативные зоны находятся рядом с соответствующими функционально сенсорными зонами. К примеру, какая-либо ассоциативная зрительная функция контролируется зрительной ассоциативной зоной, которая расположена рядом с сенсорным зрительным участком.
Установление закономерностей работы мозга, анализ его локальных нарушений и проверку его активности осуществляет наука нейропсихология, которая находится на стыке нейробиологии, психологии, психиатрии и информатики.
Особенности локализации по полям
Кора большого мозга пластична, что сказывается на переходе функций одного отдела, если произошло его нарушение, в другой. Это обусловлено тем, что анализаторы в коре имеют ядро, где происходит высшая деятельность, и периферию, которая отвечает за процессы анализа и синтеза в примитивном виде. Между ядрами анализаторов находятся элементы, которые принадлежат разным анализаторам. Если повреждение касается ядра, за его деятельность начинают отвечать периферические составляющие.

Таким образом, локализация функций, которыми обладает кора головного мозга, – понятие относительное, так как определенных границ не существует. Тем не менее, цитоархитектоника предполагает наличие 52 полей, которые сообщаются друг с другом проводящими путями:
- ассоциативными (этот тип нервных волокон отвечает за деятельность коры в области одного полушария);
- комиссуральными (связывают симметричные области обоих полушарий);
- проекционными (способствуют сообщению коры, подкорковых структур с другими органами).
Таблица 1
|
Соответствующие поля |
||
|
Двигательная |
||
|
Чувствительная |
||
|
Зрительная |
||
|
Обонятельная |
||
|
Вкусовая |
||
|
Речедвигательная, которая включает центры: |
Вернике, позволяющий воспринимать устную речь |
|
|
Брока – отвечает за движение языковых мышц; поражение грозит полной потерей речи |
||
|
Восприятия речи на письме |
||

Итак, строение коры головного мозга предполагает рассмотрение ее в горизонтальной и вертикальной ориентации. В зависимости от этого, выделяют вертикальные колонки нейронов и зоны, расположенные в горизонтальной плоскости. Основные функции, которые выполняет кора, сводятся к осуществлению поведения, регуляции мышления, сознания. Кроме того, она обеспечивает взаимодействие организма с внешней средой и принимает участие в контроле работы внутренних органов.
Головной мозг располагается в мозговом отделе черепа. Его средний вес 1360 г. Выделяют три больших отдела мозга: ствол, подкорковый отдел и кару больших полушарий. Из основания мозга выходят 12 пар черепных нервов.
1 - верхний участок спинного мозга; 2 - продолговач ый мозг, 3 - мост, 4 - мозжечок; 5 - средний мозг; 6 - четверохолмие; 7 - промежуточный мозг; 8 - кора больших полушарий; 9 - мозолистое тело, соединяющее правое полушарие с новым; 10 - перекрест зрительных нервов; 11 - обонятельные луковицы.
Отделы головного мозга и их функции
|
Отделы мозга |
Структуры отделов |
Функции |
|
|
СТВОЛ МОЗГА |
Задний мозг |
Продолговатый мозг Здесь находятся ядра с отходящими парами черепно-мозговы> нервов: XII - подъязычных; XI - добавочных; X - блуждающих; IX - языкоглоточных нервов |
Проводниковая - связь спинного и вышележащих отделов головного мозга. Рефлекторные: 1) регуляция деятельности дыхательной, сердечно-сосудистой и пищеварительной систем; 2) пищевые рефлексы слюноотделения, жевания, глотания; 3) защитные рефлексы: чихание, моргание, кашель, рвота; |
|
Варолиев мост содержит ядра: VIII - слухового; VII - лицевого; VI - отводящего; V - тройничного нервов. |
Проводниковая - содержит восходящие и нисходящие нервные пути и нервные волокна, соединяющие полушария мозжечка между собой и с корой большого мозга. Рефлекторная - отвечает за вестибулярные и шейные рефлексы, регулирующие тонус мышц, в т.ч. мимических мышц. |
||
|
Мозжечок Полушария мозжечка соединены между собой и образованы серым и белым веществом. |
Координация произвольных движений и сохранение положения тела в пространстве. Регуляция мышечного тонуса и равновесия. |
||
|
Ретикулярная формация - сеть нервных волокон, оплетающих ствол мозга и промежуточный мозг. Обеспечивает взаимодействие восходящих и нисходящих путей мозга, координацию различных функций организма и регуляцию возбудимости всех отделов ЦНС. |
|||
|
Средний мозг |
Четверохолмие С ядрами первичных зрительных и слуховых центров. Ножки мозга С ядрами IV - глазодвигательного III - блокового нервов. |
Проводниковая. Рефлекторны: 1) ориентировочные рефлексы на зрительные и звуковые раздражители,которые проявляются в повороте головы и туловища; 2) регуляция мышечного тонуса и позы тела. |
|
|
ПОДКОРКА |
Передний мозг |
Промежуточный мозг: а) таламус (зрительный бугор) с ядрами ll -й пары зрительных нервов; |
Сбор и оценка всей поступающей информации от органов чувств. Выделение и передача в кору мозга наиболее важной информации. Регуляция эмоционального поведения. |
|
б) гипоталамус. |
Высший подкорковый центр вегетативной нервной системы и всех жизненно важных функций организма. Обеспечение постоянства внутренней среды и обменных процессов организма. Регуляция мотивированного поведения и обеспечение защитных реакций (жажда, голод, насыщение, страх, ярость, удовольствие и неудовольствие). Участие в смене сна и бодрствования. |
||
|
Базальные ганглии (подкорковые ядра) |
Роль в регуляции и координации двигательной активности (вместе с таламусом и мозжечком). Участие в создании и запоминании программ целенаправленных движений,обучения и памяти. |
||
|
КОРА БОЛЬШИХ ПОЛУШАРИЙ |
Древняя и старая кора (обонятельный и висцеральный мозг) Содержит ядра 1-ой пары обонятельных нервов. |
Древняя и старая кора вместе с некоторыми подкорковыми структурами формирует лимбическую систему, которая: 1) отвечает за врожденные поведенческие акты и формирование эмоций; 2) обеспечивает гомеостаз и контроль реакций, направленных на самосохранение и сохранение вида: 3 влияет на регуляцию вегетативных функций. |
|
|
Новая кора |
1) Осуществляет высшую нервную деятельность, отвечает за сложное сознательное поведение и мышление. Развитие морали, воли, интеллекта, связаны с деятельностью коры. 2) Осуществляет восприятие, оценку и обработку всей поступающей информации от органов чувств. 3) Координирует деятельность всех систем организма. 4) Обеспечивает взаимодействие организма с внешней средой. |
||
Кора больших полушарий головного мозга
Кора больших полушарий - филогенетически наиболее молодое образование мозга. За счет борозд общая площадь поверхности коры взрослого человека 1700 2000 см2. В коре насчитывают от 12 до 18 млрд, нервных клеток, которые расположены в несколько слоев. Кора представляет собой слой серого вещества толщиной 1,5-4 мм.
На рисунке ниже показаны функциональные зоны и доли коры головного мозга

|
Расположение серого и белого вещества |
Доли полушарий |
Зоны полушарий |
|
|
Кора – серое вещество, белое вещество нахо-дится под ко-рой, в белом веществе есть скопления серо-го вещества в виде ядер |
Центры речи |
||
|
Теменная |
Кожно-мышечная зона |
Контроль дви-жений, спо-собность раз-личать раздражения |
|
|
Височная |
Слуховая зона |
Дуги рефлексов, различающих звуковые раздражения |
|
|
Вкусовая и обонятельная зоны |
Рефлексы различения вкусов и запахов |
||
|
Затылочная |
Зрительная зона |
Различение зрительных раздражений |
Чувствительная и двигательная зоны коры больших полушарий
|
Левое полушарие мозга |
Правое полушарие мозга |
|
Левое полушарие ("мыслительное”, логическое) - - отвечает за регуляцию речевой деятельности, устной речи, письма, счета и логического мышления. Доминантное у правшей. |
Правое полушарие ("художественное", эмоциональное) - - участвует в распознавании зрительных, музыкальных образов, формы и структуры предметов, в сознательной ориентации в пространстве. |
|
Поперечный срез левого полушария через чувствительные центры
Представительство тела в чувствительной зоне коры больших полушарий. Чувствительная зона каждого полушария получает информацию от мышц, кожи и внутренних органов противоположной стороны тела. Рейтинг 5.00 |

Кора головного мозга - высший отдел центральной нервной системы, обеспечивающий функционирование организма как единого целого при его взаимодействии с окружающей средой.
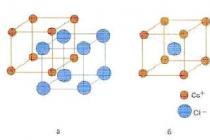
головного мозга (кора большого мозга, новая кора) представляет собой слой серого вещества, состоящего из 10-20 млрд и покрывающего большие полушария (рис. 1). Серое вещество коры составляет более половины всего серого вещества ЦНС. Суммарная площадь серого вещества коры — около 0,2 м 2 , что достигается извилистой складчатостью ее поверхности и наличием борозд разной глубины. Толщина коры в ее разных участках колеблется от 1,3 до 4,5 мм (в передней центральной извилине). Нейроны коры располагаются в шести слоях, ориентированных параллельно ее поверхности.
В участках коры, относящихся к , имеются зоны с трехслойным и пятислойным расположением нейронов в структуре серого вещества. Эти участки филогенетически древней коры занимают около 10% поверхности полушарий мозга, остальные 90% составляют новую кору.

Рис. 1. Моля латеральной поверхности коры большого мозга (по Бродману)
Строение коры головного мозга
Кора большого мозга имеет шестислойное строение
Нейроны разных слоев различаются по цитологическим признакам и функциональным свойствам.
Молекулярный слой — самый поверхностный. Представлен небольшим числом нейронов и многочисленными ветвящимися дендритами пирамидных нейронов, лежащих в более глубоких слоях.
Наружный зернистый слой сформирован плотно расположенными многочисленными мелкими нейронами разной формы. Отростки клеток этого слоя образуют кортикокортикальные связи.
Наружный пирамидальный слой состоит из пирамидных нейронов средней величины, отростки которых также участвуют в образовании кортикокортикальных связей между соседними областями коры.
Внутренний зернистый слой подобен второму слою по виду клеток и расположению волокон. В слое проходят пучки волокон, связывающие различные участки коры.
К нейронам этого слоя проводятся сигналы от специфических ядер таламуса. Слой очень хорошо представлен в сенсорных областях коры.
Внутренний пирамидный слои образован средними и крупными пирамидными нейронами. В двигательной области коры эти нейроны особенно крупные (50-100 мкм) и получили название гигантских, пирамидных клеток Беца. Аксоны этих клеток формируют быстропроводящие (до 120 м/с) волокна пирамидного тракта.
Слой полиморфных клеток представлен преимущественно клетками, аксоны которых образуют кортикоталамические пути.

Нейроны 2-го и 4-го слоев коры участвуют в восприятии, переработке поступающих к ним сигналов от нейронов ассоциативных областей коры. Сенсорные сигналы из переключающих ядер таламуса поступают преимущественно к нейронам 4-го слоя, выраженность которого наибольшая в первичных сенсорных областях коры. К нейронам 1-го и других слоев коры поступают сигналы из других ядер таламуса, базальных ганглиев, ствола мозга. Нейроны 3-го, 5-го и 6-го слоев формируют эфферентные сигналы, посылаемые в другие области коры и по нисходящим путям в нижележащие отделы ЦНС. В частности, нейроны 6-го слоя формируют волокна, следующие в таламус.
В нейронном составе и цитологических особенностях разных участков коры имеются значительные отличия. По этим отличиям Бродман разделил кору на 53 цитоархитектонических поля (см. рис. 1).
Расположение многих из этих нолей, выделенных на основе гистологических данных, совпадает по топографии с расположением корковых центров, выделенных на основе выполняемых ими функций. Используются и другие подходы деления коры на области, например, на основе содержания в нейронах определенных маркеров, по характеру нейронной активности и другим критериям.
Белое вещество полушарий головного мозга образовано нервными волокнами. Выделяют ассоциативные волокна, подразделяемые на дугообразные волокна, но которым сигналы передаются между нейронами рядом лежащих извилин и длинные продольные пучки волокон, доставляющие сигналы к нейронам более удаленных участков одноименного полушария.
Комиссуральные волокна - поперечные волокна, передающие сигналы между нейронами левого и правого полушарий.
Проекционные волокна - проводят сигналы между нейронами коры и других отделов мозга.
Перечисленные виды волокон участвуют в создании нейронных цепей и сетей, нейроны которых расположены на значительных расстояниях друг от друга. В коре имеется также особый вид локальных нейронных цепей, образованных рядом расположенными нейронами. Эти нейронные структуры получили название функциональных кортикальных колонок. Нейронные колонки образованы группами нейронов, расположенных друг над другом перпендикулярно поверхности коры. Принадлежность нейронов к одной и той же колонке можно определить по повышению их электрической активности на раздражение одного и того же рецептивного поля. Такая активность регистрируется при медленном перемещении регистрирующего электрода в коре в перпендикулярном направлении. Если регистрировать электрическую активность нейронов, расположенных в горизонтальной плоскости коры, то отмечается повышение их активности при раздражении различных рецептивных полей.
Диаметр функциональной колонки составляет до 1 мм. К нейронам одной функциональной колонки поступают сигналы от одного и того же афферентного таламокортикального волокна. Нейроны соседних колонок связаны друг с другом отростками, с помощью которых обмениваются информацией. Наличие в коре таких взаимосвязанных функциональных колонок увеличивает надежность восприятия и анализа информации, поступающей к коре.
Эффективность восприятия, обработки и использования информации корой для регуляции физиологических процессов обеспечивается также соматотопическим принципом организации сенсорных и моторных полей коры. Суть такой организации заключается в том, что в определенной (проекционной) области коры представлены не любые, а топографически очерченные участки рецептивного поля поверхности тела, мышц, суставов или внутренних органов. Так, например, в соматосенсорной коре поверхность тела человека спроецирована в виде схемы, когда в определенной точке коры представлены рецептивные поля конкретной области поверхности тела. Строгим топографическим образом в первичной моторной коре представлены эфферентные нейроны, активация которых вызывает сокращение определенных мышц тела.
Полям коры присущ также экранный принцип функционирования. При этом рецепторный нейрон посылает сигнал не на одиночный нейрон или в одиночную точку коркового центра, а на сеть или ноле нейронов, связанных отростками. Функциональными ячейками этого поля (экрана) являются колонки нейронов.
Кора мозга, формируясь на поздних этапах эволюционного развития высших организмов, в определенной мере подчинила себе все нижележащие отделы ЦНС и способна корригировать их функции. В то же время функциональная активность коры больших полушарий определяется притоком к ней сигналов от нейронов ретикулярной формации ствола мозга и сигналов от рецептивных полей сенсорных систем организма.
Функциональные области коры мозга
По функциональному признаку в коре выделяют сенсорные, ассоциативные и двигательные области.
Сенсорные (чувствительные, проекционные) области коры
Они состоят из зон, содержащих нейроны, активация которых афферентными импульсами от сенсорных рецепторов или прямым воздействием раздражителей вызывает появление специфических ощущений. Эти зоны имеются в затылочной (поля 17-19), теменной (ноля 1-3) и височной (поля 21-22, 41-42) областях коры.
В сенсорных зонах коры выделяют центральные проекционные поля, обеспечивающие топкое, четкое восприятие ощущений определенных модальностей (свет, звук, прикосновение, тепло, холод) и вторичные проекционные ноля. Функцией последних является обеспечение понимания связи первичного ощущения с другими предметами и явлениями окружающего мира.
Зоны представительства рецептивных полей в сенсорных зонах коры в значительной мере перекрываются. Особенность нервных центров в области вторичных проекционных полей коры — их пластичность, которая проявляется возможностью перестройки специализации и восстановления функций после повреждения какого-либо из центров. Эти компенсаторные возможности нервных центров особенно выражены в детском возрасте. В то же время повреждение центральных проекционных полей после перенесенных заболевании, сопровождается грубым нарушением функций чувствительности и часто невозможностью ее восстановления.
Зрительная кора
Первичная зрительная кора (VI, поле 17) располагается по обеим сторонам шпорной борозды на медиальной поверхности затылочной доли головного мозга. В соответствии с выявлением па неокрашенных срезах зрительной коры чередующихся белых и темных полос ее называют также стриарной (полосатой) корой. К нейронам первичной зрительной коры посылают зрительные сигналы нейроны латерального коленчатого тела, которые получают сигналы от ганглиозных клеток сетчатки. Зрительная кора каждого полушария получает визуальные сигналы от ипсилатеральной и контралатеральной половин сетчатки обоих глаз и их поступление к нейронам коры организовано по соматотопическому принципу. Нейроны, к которым поступают зрительные сигналы от фоторецепторов, топографически расположены в зрительной коре подобно рецепторам в сетчатке глаза. При этом область желтого пятна сетчатки имеет относительно большую зону представительства в коре, чем другие области сетчатки.
Нейроны первичной зрительной коры ответственны за зрительное восприятие, которое на основе анализа входных сигналов проявляется их способностью обнаруживать зрительный стимул, определять его специфическую форму и ориентацию в пространстве. Упрощенно можно представить сенсорную функцию зрительной коры в решении задачи и ответе на вопрос, что представляет собой зрительный объект.
В анализе других качеств зрительных сигналов (например, расположения в пространстве, движения, связи с другими событиями и т.д.) принимают участие нейроны полей 18 и 19 экстрастриарной коры, расположенных но соседству с нолем 17. Информация о сигналах, поступивших в сенсорные зрительные зоны коры, передастся для дальнейшего анализа и использования зрения для выполнения других функций мозга в ассоциативные области коры и другие отделы мозга.
Слуховая кора
Расположена в латеральной борозде височной доли в области извилины Гешля (AI, поля 41-42). К нейронам первичной слуховой коры поступают сигналы от нейронов медиальных коленчатых тел. Волокна слуховых путей, проводящие звуковые сигналы в слуховую кору, организованы тонотопически, и это позволяет нейронам коры получать сигналы от определенных слуховых рецепторных клеток кортиева органа. Слуховая кора регулирует чувствительность слуховых клеток.
В первичной слуховой коре формируются звуковые ощущения и проводится анализ отдельных качеств звуков, позволяющий ответить на вопрос, что представляет собой воспринятый звук. Первичная слуховая кора играет важную роль в анализе коротких звуков, интервалов между звуковыми сигналами, ритма, звуковой последовательности. Более сложный анализ звуков осуществляется в ассоциативных областях коры, смежных с первичной слуховой. На основе взаимодействия нейронов этих областей коры осуществляется бинауральный слух, определяются характеристики высоты, тембра, громкости звука, принадлежность звука, формируется представление о трехмерном звуковом пространстве.
Вестибулярная кора
Располагается в верхней и средней височных извилинах (поля 21-22). К ее нейронам поступают сигналы от нейронов вестибулярных ядер ствола мозга, связанных афферентными связями с рецепторами полукружных каналов вестибулярного аппарата. В вестибулярной коре формируется ощущение о положении тела в пространстве и ускорении движений. Вестибулярная кора взаимодействует с мозжечком (через височно-мостомозжечковый путь), участвует в регуляции равновесия тела, приспособлении позы к осуществлению целенаправленных движений. На основе взаимодействия этой области с соматосенсорной и ассоциативными областями коры происходит осознание схемы тела.
Обонятельная кора
Расположена в области верхней части височной доли (крючок, ноля 34, 28). Кора включает ряд ядер и относится к структурам лимбической системы. Ее нейроны расположены в трех слоях и получают афферентные сигналы от митральных клеток обонятельной луковицы, связанных афферентными связям с обонятельными рецепторными нейронами. В обонятельной коре проводится первичный качественный анализ запахов и формируется субъективное ощущение запаха, его интенсивности, принадлежности. Повреждение коры ведет к снижению обоняния или к развитию аносмии — потере обоняния. При искусственном раздражении этой области возникают ощущения различных запахов по типу галлюцинаций.
Вкусовая кора
Расположена в нижней части соматосенсорной извилины, непосредственно кпереди от области проекции лица (поле 43). Ее нейроны получают афферентные сигналы от релейных нейронов таламуса, которые связаны с нейронами ядра одиночного тракта продолговатого мозга. К нейронам этого ядра поступают сигналы непосредственно от чувствительных нейронов, образующих синапсы на клетках вкусовых луковиц. Во вкусовой коре проводится первичный анализ вкусовых качеств горького, соленого, кислого, сладкого и на основе их суммации формируется субъективное ощущение вкуса, его интенсивности, принадлежности.
Сигналы запахов и вкуса достигают нейронов передней части островковой коры, где на основе их интеграции формируется новое, более сложное качество ощущений, определяющее наше отношение к источникам запаха или вкуса (например, к пище).
Соматосенсорная кора
Занимает область постцентральной извилины (SI, поля 1-3), включая парацентральную дольку на медиальной стороне полушарий (рис. 9.14). В соматосенсорную область поступают сенсорные сигналы от нейронов таламуса, связанных спиноталамическими путями с рецепторами кожи (тактильная, температурная, болевая чувствительность), проприорецепторами (мышечных веретен, суставных сумок, сухожилий) и интерорецепторами (внутренних органов).

Рис. 9.14. Важнейшие центры и области коры большого мозга
Из-за перекреста афферентных путей в соматосенсорную зону левого полушария приходит сигнализация от правой стороны тела, соответственно в правое полушарие — от левой стороны тела. В этой сенсорной области коры соматотопически представлены все части тела, но при этом наиболее важные рецептивные зоны пальцев рук, губ, кожи лица, языка, гортани занимают относительно большие площади, чем проекции таких поверхностей тела, как спина, передняя часть туловища, ноги.
Расположение представительства чувствительности частей тела вдоль постцентральной извилины часто называют «перевернутый гомункулюс», так как проекция головы и шеи находится в нижней части постцентральной извилины, а проекция каудальной части туловища и ног — в верхней части. При этом чувствительность голеней и стоп проецируется на кору пара- центральной дольки медиальной поверхности полушарий. Внутри первичной соматосенсорной коры имеется определенная специализация нейронов. Например, нейроны поля 3 получают преимущественно сигналы от мышечных веретен и механорецепторов кожи, поля 2 — от рецепторов суставов.
Кору постцентральной извилины относят к первичной соматосенсорной области (SI). Ее нейроны посылают обработанные сигналы к нейронам вторичной соматосенсорной коры (SII). Она располагается кзади от постцентральной извилины в теменной коре (поля 5 и 7) и принадлежит к ассоциативной коре. Нейроны SII не получают прямых афферентных сигналов от нейронов таламуса. Они связаны с нейронами SI и нейронами других областей коры мозга. Это позволяет проводить здесь интегральную оценку сигналов, попадающих в кору по спиноталамическому пути с сигналами, поступающими из других (зрительной, слуховой, вестибулярной и т.д.) сенсорных систем. Важнейшей функцией этих полей теменной коры является восприятие пространства и трансформация сенсорных сигналов в координаты моторных. В теменной коре формируется стремление (намерение, побуждение) осуществить моторное действие, что является основой для начала планирования в ней предстоящей моторной активности.

Интеграция различных сенсорных сигналов связана с формированием различных ощущений, адресуемых к разным частям тела. Эти ощущения используются как для формирования психических, так и других ответных реакций, примерами которых могут быть движения при одновременном участии мышц обеих сторон тела (например, перемещение, ощупывание обеими руками, хватание, однонаправленное движение обеими руками). Функционирование этой области необходимо для узнавания предметов на ощупь и определения пространственного расположения этих предметов.
Нормальная функция соматосенсорных областей коры является важным условием формирования таких ощущений как тепло, холод, боль и их адресации к определенной части тела.
Повреждение нейронов области первичной соматосенсорной коры ведет к снижению различных видов чувствительности на противоположной стороне тела, а локальное повреждение — к потере чувствительности в определенной части тела. Особенно ранимой при повреждении нейронов первичной соматосенсорной коры является дискриминационная чувствительность кожи, а наименее — болевая. Повреждение нейронов вторичной соматосенсорной области коры может сопровождаться нарушением способности распознания предметов на ощупь (тактильная агнозия) и навыков использования предметов (апраксия).
Двигательные области коры
Около 130 лет тому назад исследователи, нанося точечные раздражения на кору мозга электрическим током, обнаружили, что воздействие на поверхность передней центральной извилины вызывает сокращение мышц противоположной стороны тела. Так было обнаружено наличие одной из моторных зон коры мозга. В последующем оказалось, что к организации движений имеют отношение несколько областей коры мозга и его другие структуры, а в областях моторной коры имеются не только двигательные нейроны, но и нейроны, осуществляющие другие функции.
Первичная моторная кораПервичная моторная кора располагается в передней центральной извилине (MI, поле 4). Ее нейроны получают основные афферентные сигналы от нейронов соматосенсорной коры — полей 1, 2, 5, премоторной коры и таламуса. Кроме того, через вентролатеральный таламус в MI посылают сигналы нейроны мозжечка.
От пирамидных нейронов Ml начинаются эфферентные волокна пирамидного пути. Часть волокон этого пути следует к моторным нейронам ядер черепных нервов ствола мозга (кортикобульбарный тракт), часть — к нейронам стволовых моторных ядер (красное ядро, ядра ретикулярной формации, стволовые ядра, связанные с мозжечком) и часть — к интер- и моторным нейронам спинного мозга (кортикоспинальный тракт).
Имеется соматотопическая организация расположения нейронов в MI, контролирующих сокращение разных мышечных групп тела. Нейроны, контролирующие мышцы ног и туловища, расположены в верхних участках извилины и занимают относительно малую площадь, а контролирующие мышцы рук, особенно пальцев, лица, языка и глотки расположены в нижних участках и занимают большую площадь. Таким образом, в первичной двигательной коре относительно большую площадь занимают те нейронные группы, которые управляют мышцами, осуществляющими разнообразные, точные, мелкие, тонко регулируемые движения.
Поскольку многие нейроны Ml увеличивают электрическую активность непосредственно перед началом произвольных сокращений, то первичной моторной коре отводят ведущую роль в контроле активности моторных ядер ствола и мотонейронов спинного мозга и инициации произвольных, целенаправленных движений. Повреждение поля Ml ведет к парезу мышц и невозможности осуществления тонких произвольных движений.
Вторичная моторная кораВключает области премоторной и дополнительной моторной коры (МII, поле 6). Премоторная кора расположена в поле 6, на боковой поверхности мозга, кпереди от первичной моторной коры. Ее нейроны получают через таламус афферентные сигналы из затылочной, соматосенсорной, теменной ассоциативной, префронтальной областей коры и мозжечка. Обработанные в ней сигналы нейроны коры посылают по эфферентным волокнам в моторную кору MI, небольшое число — в спинной мозг и большее — в красные ядра, ядра ретикулярной формации, базальные ганглии и мозжечок. Премоторная кора играет основную роль в программировании и организации движений, находящихся под контролем зрения. Кора участвует в организации позы и вспомогательных движений для действий, осуществляемых дистальными мышцами конечностей. Повреждение прсмотор- ной коры часто вызывает тенденцию повторного выполнения начатого движения (персеверация), даже если осуществленное движение достигло цели.
В нижней части премоторной коры левой лобной доли, непосредственно кпереди от участка первичной моторной коры, в которой представлены нейроны, контролирующие мышцы лица, располагается речевая область , или моторный центр речи Брока. Нарушение ее функции сопровождается нарушением артикуляции речи, или моторной афазией.
Дополнительная моторная кора располагается в верхней части поля 6. Ее нейроны получают афферентные сигналы из соматосснсорной, теменной и префронтальной областей коры головного мозга. Обработанные в ней сигналы нейроны коры посылают по эфферентным волокнам в первичную моторную кору MI, спинной мозг, стволовые моторные ядра. Активность нейронов дополнительной моторной коры повышается раньше, чем нейронов коры MIи главным образом в связи с осуществлением сложных движений. При этом возрастание нейронной активности в дополнительной моторной коре не связано с движениями как таковыми, для этого достаточно мысленно представить модель предстоящих сложных движений. Дополнительная моторная кора принимает участие в формировании программы предстоящих сложных движений и в организации моторных реакций на специфичность сенсорных стимулов.
Поскольку нейроны вторичной моторной коры посылают множество аксонов в поле MI, ее считают в иерархии моторных центров организации движений более высокой структурой, стоящей над моторными центрами моторной коры MI. Нервные центры вторичной моторной коры могут оказывать влияние на активность моторных нейронов спинного мозга двумя путями: непосредственно через кортикоспинальный путь и через поле MI. Поэтому их иногда называют супрамоторными полями, в функцию которых входит инструктирование центров поля MI.
Из клинических наблюдений известно, что сохранение нормальной функции вторичной моторной коры важно для осуществления точных движений руки, и особенно для выполнения ритмических движений. Так, например, при их повреждении пианист перестает чувствовать ритм и выдерживать интервал. Нарушается способность к осуществлению противоположных движений руками (манипулирование обоими руками).
При одновременном повреждении моторных зон MI и MII коры утрачивается способность к тонким координированным движениям. Точечные раздражения в этих областях моторной зоны сопровождаются активацией не отдельных мышц, а целой группы мышц, вызывающих направленное движение в суставах. Эти наблюдения послужили поводом для формирования вывода о том, что в моторной коре представлены не столько мышцы, сколько движения.
Префронтальная кораРасполагается в области поля 8. Ее нейроны получают основные афферентные сигналы из затылочной зрительной, теменной ассоциативной коры, верхних холмиков четверохолмия. Обработанные сигналы передаются по эфферентным волокнам в премоторную кору, верхние холмики четверохолмия, стволовые моторные центры. Кора играет определяющую роль в организации движений, находящихся под контролем зрения и принимает непосредственное участие в инициации и контроле движений глаз и головы.
Механизмы, реализующие превращение замысла движения в конкретную моторную программу, в залпы импульсов, посылаемых к определенным мышечным группам, остаются недостаточно понятными. Считается, что замысел движения формируется благодаря функциям ассоциативной и других областей коры, взаимодействующих со многими структурами головного мозга.
Информация о замысле движения передается в двигательные области лобной коры. Двигательная кора через нисходящие пути активирует системы, обеспечивающие выработку и использование новых двигательных программ или использование старых, уже отработанных на практике и хранящихся в памяти. Составной частью этих систем являются базальные ганглии и мозжечок (см. их функции выше). Программы движения, выработанные при участии мозжечка и базальных ганглиев, передаются через таламус в моторные зоны и прежде всего в первичную моторную область коры. Эта область непосредственно инициирует исполнение движений, подключая к нему определенные мышцы и обеспечивая последовательность смены их сокращения и расслабления. Команды коры передаются на моторные центры ствола мозга, спинальные мотонейроны и мотонейроны ядер черепных нервов. Мотонейроны в осуществлении движений выполняют роль конечного пути, через который двигательные команды передаются непосредственно к мышцам. Особенности передачи сигналов от коры к моторным центрам ствола и спинного мозга описаны в главе, посвященной ЦНС (ствол мозга, спинной мозг).
Ассоциативные области коры
У человека ассоциативные области коры занимают около 50% площади всей коры большого мозга. Они располагаются в участках между сенсорными и двигательными областями коры. Ассоциативные области не имеют четких границ со вторичными сенсорными областями как по морфологическим, так и по функциональным признакам. Выделяют теменную, височную и лобную ассоциативные области коры больших полушарий.
Теменная ассоциативная область коры. Располагается в полях 5 и 7 верхней и нижней теменных долек мозга. Область граничит впереди с соматосенсорной корой, сзади — со зрительной и слуховой корой. К нейронам теменной ассоциативной области могут поступать и активировать их зрительные, звуковые, тактильные, проприоцептивные, болевые, сигналы из аппарата памяти и другие сигналы. Часть нейронов является полисенсорной и может повышать свою активность при поступлении к ней соматосенсорных и визуальных сигналов. Однако степень повышения активности нейронов ассоциативной коры на поступление афферентных сигналов зависит от текущей мотивации, внимания субъекта и информации, извлекаемой из памяти. Она остается незначительной, если поступающий из сенсорных областей мозга сигнал для субъекта безразличен, и существенно возрастает, если он совпал с имеющейся мотивацией и привлек его внимание. Например, при предъявлении обезьяне банана активность нейронов ассоциативной теменной коры остается невысокой, если животное сыто, и наоборот, активность резко возрастает у голодных животных, которым нравятся бананы.
Нейроны теменной ассоциативной коры связаны эфферентными связями с нейронами префронтальной, премоторной, моторной областей лобной доли и поясной извилины. Исходя из экспериментальных и клинических наблюдений, принято считать, что одной из функций коры поля 5 является использование соматосенсорной информации для осуществления целенаправленных произвольных движений и манипулирования объектами. Функцией коры поля 7 является интеграция визуальных и соматосенсорных сигналов для координации движений глаз и визуально-ведомых движений руки.
Нарушением этих функций теменной ассоциативной коры при повреждении ее связей с корой лобной доли или заболеванием самой лобной доли, объясняются симптомы последствий заболеваний, локализованных в области теменной ассоциативной коры. Они могут проявляться затруднением в понимании смыслового содержания сигналов (агнозия), примером которого может быть потеря способности распознавания формы и пространственного расположения объекта. Могут нарушаться процессы трансформации сенсорных сигналов в адекватные моторные действия. В последнем случае больной теряет навыки практического использования хорошо знакомых инструментов и предметов (апраксия), и у него может развиться невозможность осуществления визуально-ведомых движений (например, движение руки в направлении предмета).
Лобная ассоциативная область коры. Располагается в префронтальной коре, которая является частью коры лобной доли, локализующейся кпереди от полей 6 и 8. Нейроны лобной ассоциативной коры получают обработанные сенсорные сигналы по афферентным связям от нейронов коры затылочной, теменной, височной долей мозга и от нейронов поясной извилины. Лобная ассоциативная кора получает сигналы о текущем мотивационном и эмоциональном состояниях от ядер таламуса, лимбической и других структур мозга. Кроме того, лобная кора может оперировать абстрактными, виртуальными сигналами. Эфферентные сигналы ассоциативная лобная кора посылает обратно, в структуры мозга, от которых они были получены, в моторные области лобной коры, хвостатое ядро базальных ганглиев и гипоталамус.
Эта область коры играет первостепенную роль в формировании высших психических функций человека. Она обеспечивает формирование целевых установок и программ осознанных поведенческих реакций, узнавание и смысловую оценку предметов и явлений, понимание речи, логическое мышление. После обширных повреждений лобной коры у больных могут развиться апатия, снижение эмоционального фона, критичного отношения к своим собственным поступкам и поступкам окружающих, самодовольство, нарушение возможности использования прошлого опыта для изменения поведения. Поведение больных может стать непредсказуемым и неадекватным.
Височная ассоциативная область коры. Располагается в полях 20, 21, 22. Нейроны коры получают сенсорные сигналы от нейронов слуховой, экстрастриарной зрительной и префронтальной коры, гиппокампа и миндалины.
После двухстороннего заболевания височных ассоциативных областей с вовлечением в патологический процесс гиппокампа или связей с ним у больных могут развиться выраженные нарушения памяти, эмоционального поведения, неспособность сосредоточения внимания (рассеянность). У части людей при повреждении нижневисочной области, где предположительно располагается центр узнавания лица, может развиться зрительная агнозия — неспособность узнавания лиц знакомых людей, предметов, при сохранности зрения.
На границе височной, зрительной и теменной областей коры в нижней теменной и задней части височной доли располагается ассоциативный участок коры, получивший название сенсорного центра речи, или центра Вернике. После его повреждения развивается нарушение функции понимания речи при сохранности речедвигательной функции.
Локализация функций в больших полушариях. Кора больших полушарий головного мозга делится на основные зоны, состоящие из нескольких корковых полей. Каждая из этих зон выполняет определенную общую функцию, а составляющие ее поля специализированно участвуют в реализации отдельных элементов этой функции. Однако благодаря проводящим путям в осуществлении отдельных звеньев высшей и низшей нервной деятельности участвует несколько зон больших полушарий, определенные подкорковые центры, ядра мозгового ствола и сегменты спинного мозга.
При тонкой и точной специализации определенных групп нейронов головной и спинной мозг функционируют как единое целое. Психические функции головного мозга также не ограничены отдельными участками коры, а являются результатом совместной деятельности обширных зон больших полушарий и подкорковых центров.
Рис. 123. Индивидуальные изменения основных полей новой коры больших полушарий у трех взрослых (А, Б, В). Цифры- поля по Бродману
Моторная зона (поле 4) расположена в передней центральной извилине вдоль центральной борозды. В верхней четверти зоны находятся двигательные центры для мышц ног.
Сверху расположены нейроны, иннервирующие мышцы пальцев ног, а снизу - бедра и туловища. Две средние четверти заняты центрами для рук, выше - центр мышц лопатки, а ниже - мышц пальцев. И, наконец, в нижней четверти передней центральной извилины находятся центры мышц лица и речевого аппарата.
В результате исторического развития головного мозга человека в процессе труда и речи особенно большое место занимают группы нейронов, которые вызывают сокращение мышц кисти руки, главным образом большого пальца, и мышц лица, языка и гортани. К ним поступают центростремительные волокна из проприорецепторов, входящие по задним корешкам в спинной мозг, где они поднимаются в составе заднего столба той же стороны до ядер нежного и клиновидного пучков продолговатого мозга. Из этих ядер выходят волокна вторых нейронов, образующие медиальную петлю и после перекреста достигающие ядер зрительного бугра противоположной стороны. Отсюда большая часть центростремительных волокон третьих нейронов достигает задней центральной извилины и далее поступает в переднюю центральную извилину, а меньшая часть входит в нее прямым путем. Таким образом, передняя центральная извилина посредством волокон, проходящих в проводящих путях коры, связана с задней центральной извилиной. Из моторной зоны выходят центробежные двигательные волокна пирамидных нейронов, которые составляют пирамидные проводящие пути; они достигают нейронов передних рогов спинного мозга. Моторная зона вызывает координированные движения скелетных мыщц, преимущественно на противоположной стороне тела. Она функционирует совместно с подкорковыми центрами - полосатыми телами, а также люисовым телом, красным ядром и черным веществом.
При поражениях определенных участков передней центральной извилины нарушаются произвольные движения отдельных групп мышц. Неполное поражение зоны вызывает нарушение движений- парез, а полное ее разрушение - паралич.
Зона кожно-мышечной чувствительности (поля 1, 2, 3, 43 и частично 5 и 7) расположена в задней центральной извилине вдоль задней центральной борозды. В этой зоне особенно сильно развиты зернистые слои коры, к которым подходят центростремительные волокна из рецепторов кожи, идущие в составе тех же проводящих путей, как и волокна из проприорецепторов. Расположение воспринимающих групп нейронов такое же, как в моторной зоне. Наибольшую поверхность занимают нейроны, воспринимающие импульсы из рецепторов кисти руки, лица, языка и гортани. Поле 7 больше других полей связано с чувствительностью руки. Зона кожно-мышечной чувствительности не полностью отграничена от моторной зоны, так как в полях 3, 4 и 5 происходит сочетание зернистых нейронов с гигантскими пирамидными нейронами. В моторной зоне находится примерно 80% двигательных нейронов, а в зоне кожно-мышечной чувствительности - 20%. В каждое полушарие поступают импульсы главным образом из рецепторов противоположной стороны тела, но также и из рецепторов той же стороны. В эту зону поступают центростремительные импульсы преимущественно из бокового и полулунного ядер зрительного бугра.
При поражениях определенных участков задней центральной извилины нарушается чувствительность в отдельных участках кожи. Потеря способности узнавать предметы при их осязании обозначается как тактильная агнозия. При нарушениях функций зоны наблюдаются расстройства осязания, болевых и температурных ощущений кожи и мышечно-суставной чувствительности. Неполное поражение зоны вызывает понижение рецепции - гипостезию, а полное - ее потерю - анестезию.
Лобная зона (поля 6, 5, 9, 10, 11, 44, 45, 46, 47) расположена в лобной доле впереди моторной. Она делится на премоторную и речедвигательную. Премоторная зона (поля 6, 8, 9, 10, 11) регулирует тонус скелетных мышц и координированные движения тела, ориентирующие его в пространстве. С полем 10, которое участвует в выполнении двигательных условных рефлексов, функционально связано поле 46. В премоторную зону поступают центростремительные импульсы из внутренних органов и из нее исходит значительная часть центробежных вегетативных волокон. Поэтому поражение премоторной зоны вызывает нарушение координации движений - атаксию и расстройства функций сердечнососудистой, дыхательной, пищеварительной и других систем внутренних органов.
Зрительная зона (поля 17, 18, 19) расположена на внутренней поверхности затылочной доли по обеим сторонам шпорной борозды. У человека она занимает 12% общей поверхности коры. Поле 17 находится на затылочном полюсе; оно окружено полем 18, которое окружает поле 19, граничащее с задним отделом лимбической области, верхней и нижней теменными областями. В поле 17 - центральном поле зрительной зоны в 16 раз больше нейронов, чем в центральном поле слуховой зоны (поле 41), и в 10 раз больше нейронов, чем в центральном поле моторной зоны (поле 4). Это указывает на ведущее в историческом и индивидуальном развитии человека значение зрения.
Из сетчатки 900 тыс.- 1 млн. центростремительных волокон зрительных нервов доходит до наружного коленчатого тела, в котором точно проецируются отдельные части сетчатки. Центростремительные волокна нейронов наружного коленчатого тела направляются в зрительную зону, преимущественно в основное зрительное поле 17. Другими промежуточными зрительными центрами, участвующими в передаче не зрительных импульсов, а глазодвигательных, являются подушка зрительного бугра и передние бугры четверохолмия.
До поступления в наружное коленчатое тело волокна зрительного нерва перекрещиваются. Благодаря этому перекресту в составе зрительного пути, направляющегося в зрительную зону каждого полушария, 50% волокон своей стороны и 50% волокон противоположной стороны. В зрительную зону левого полушария поступают зрительные импульсы из левых половин сетчаток обоих глаз, а в зону правого полушария - из правых половин сетчаток обоих глаз. Поэтому разрушение одной из зрительных зон вызывает слепоту в одноименных половинах сетчаток в обоих глазах - гемианопсию. В зрительных нервах, кроме центростремительных волокон, проходят и несколько более толстые центробежные волокна к мышцам радужной оболочки и центробежные тонкие симпатические волокна из нейронов подкорковых центров. Небольшая часть центростремительных волокон зрительного нерва не прерывается в подкорковых образованиях, а прямо направляется в мозжечок и зрительные зоны больших полушарий.
Разрушение обоих полей 17 вызывает полную корковую слепоту, разрушение поля 18 приводит к потере зрительной памяти при сохранении зрения, что обозначается как зрительная агнозия, а разрушение поля 19 - к потере ориентации в непривычной обстановке.
Слуховая зона (поля 41, 42, 21, 22, 20, 37) расположена на поверхности височной доли, преимущественно передней поперечной височной извилины и верхней височной извилины. Поле 41, расположенное в верхней височной извилине и в передней части поперечной извилины, является проекцией кортиева органа улитки. Из органа Корти центростремительные импульсы проходят через спиральный узел по улиточному нерву, состоящему примерно из 30 тыс. волокон. В этом узле находятся первые биполярные нейроны слухового пути. Далее волокна первых нейронов передают слуховые импульсы в ядра слухового нерва в продолговатом мозге, где находятся вторые нейроны. Волокна ядер слухового нерва связываются с ядрами лицевого нерва в продолговатом мозге и глазодвигательного нерва в передних буграх среднего мозга. Поэтому при сильных звуках рефлекторно сокращаются мышцы лица, век, ушной раковины и вызываются движения глаз.
Большая часть волокон ядер слухового нерва перекрещивается в варолиевом мосту, а меньшая проходит на своей стороне. Затем волокна слухового пути поступают в боковую лемнисковую петлю, которая заканчивается в задних буграх четверохолмия и во внутреннем коленчатом теле, где находятся третьи нейроны - их волокна проводят центростремительные импульсы в слуховую зону. Существуют также прямые пути, связывающие ядра слуховых нервов с мозжечком и слуховой зоной. Большая часть прямых мозжечковых путей образуется вестибулярным нервом, а меньшая- улитковым нервом, составляющими вместе общий ствол слухового нерва. Вестибулярный аппарат проецируется также в слуховой зоне.
Разрушение поля 41 на одной стороне вызывает глухоту на противоположной стороне и ослабление слуха на своей стороне, а разрушение полей 41 на обеих сторонах ведет к полной корковой глухоте. Разрушение поля 22 в передней трети верхней височной извилины приводит к музыкальной глухоте - теряется восприятие интенсивности тона, тембра и ритма звуков - слуховая агнозия. Разрушение полей 21 и 20 в средней и нижней височных извилинах вызывает атаксию - расстройство равновесия и координации движений.
В слуховой зоне расположен также рече-слуховой центр.
Обонятельная и вкусовая зоны. Обонятельная зона находится в древней коре, в которую поступают центростремительные импульсы из обонятельных клеток. Кроме обонятельной функции, она выполняет также вкусовую и участвует в деятельности пищеварительной, выделительной и половой систем. Раньше считали, что гиппокамп выполняет обонятельную функцию. В настоящее время полагают, что вместе с лимбической системой, гипоталамической областью промежуточного мозга и гипофизом, средним и продолговатым мозгом и особенно ретикулярной формацией гиппокамп участвует в общих двигательных реакциях и вегетативных рефлексах при эмоциях. Собственно вкусовая зона, вероятно, расположена в поле 43, которое находится в нижнем отделе задней центральной извилины.
Лимбическая извилина (заднее поле 23 и переднее поле 24) и кора островка (поля 13 и 14) участвуют в высшей нервной деятельности.
Все зоны коры не обособлены, а связаны между собой проводящими путями.
Центры речи (поля 44, 45, 46, 39, 40, 42, 22,37). Двигательный центр речи расположен в нижней части передней центральной извилины в поле 44. У большинства правшей площадь поля 44 в левом полушарии больше, чем в правом полушарии. Поле 44 вызывает сложные сокращения речевой мускулатуры, необходимые для произнесения слов. При разрушении этого поля человек не может говорить, но может производить простейшие сокращения речевой мускулатуры - кричать и петь. Это моторная, двигательная афазия, которая в некоторых случаях проявляется в отсутствии сокращений мышц языка и остальной речевой мускулатуры. Так как в этих случаях слуховой центр речи не поврежден, то понимание речи окружающих сохраняется. При поражении поля 44 часто нарушается не только устная речь, но и внутренняя речь или способность формулировать мысли словами без их произнесения, на основе накопленных звуковых образов, имеющих определенное смысловое содержание. При этом затруднено чтение про себя, расстроена способность писать произвольно и под диктовку, но сохранено копирование букв при письме. У правшей моторная афазия наблюдается при поражении левого полушария, а левшей - правого.

Рис. 129. Локализация центров речи:
1 - двигательный, 2 - слуховой, 3 - зрительный
Впереди поля 44 расположено поле 45, которое регулирует построение грамматически правильных сочетаний слов и пение. При поражении этого поля вследствие потери памяти на приемы произношения пение расстраивается. Мимика и жестикуляция, придающие речи ее выразительность, осуществляются благодаря импульсам, поступающим из поля 46 в поля 44 и 45, в поля премоторной области и в подкорковые центры.
Слуховой, или сенсорный, центр речи расположен в заднем отделе левой верхней височной извилины в поле 42, которое осуществляет понимание слова при слышании его. Если поле разрушается, теряется способность понимания смысла слов, но сохраняется их восприятие как звуков - сенсорная афазия, или речевая глухота. При этом вследствие отсутствия понимания собственной речи, иногда наблюдается чрезмерная говорливость - логоррея, или словесный понос. В задней части поля 22 фиксируются связи звуковых образов слов со всеми воспринимающими зонами, в которых возникают представления о предметах и явлениях. Поэтому поражение этого поля также вызывает сенсорную афазию.
Поля 39 и 40, расположенные в теменной доле рядом с полем 22, осуществляют понимание смысла сочетаний слов или фраз. Поэтому их поражение приводит к расстройству речи, которое называется семантической (смысловой) афазией. При поражении поля 39, вследствие потери способности узнавать буквы и цифры и понимать смысл видимых письменных образов слов и цифр, теряется способность читать вслух, писать и считать. Поражение поля 40 вызывает потерю способности писать, так как отсутствует ориентация движений в пространстве и нарушена их последовательность. Это отсутствие способности производить системные, целенаправленные движения (апраксия) не исключает возможности правильно совершать отдельные движения руки, не связанные с письмом. Следовательно, процесс письма у правшей осуществляется, височной, нижнетеменной и нижнелобной областями левого полушария. При поражении поля 37 вызывается потеря памяти на слова - амнестическая афазия.
Таким образом, в осуществлении функции речи участвуют большие полушария головного мозга в целом, но особенная роль выполняется отдельными полями коры. У правшей в результате преимущественного развития функций правой руки и правой половины тела особенно развиты сложнейшие психические функции левого полушария головного мозга.
Похожие материалы: